Сушка биошлама: схемы проектирования систем для липкой фазы — химическая инженерия
| Yen-Hsuing Kiang, Xiamen Baipeng Environmental Engineering Co.
Сушка биошлама с очистных сооружений осложняется «липкой фазой», что делает эту практику скорее искусством, чем наукой. Здесь обсуждаются попытки определить начало и конец этой фазы аналитически
Сушка биошлама с очистных сооружений (СОСВ) – непростая задача. Наиболее неприятную проблему при сушке биошлама вызывает так называемая липкая фаза биошлама. Эта фаза представляет собой частично обезвоженный пастообразный материал, который прилипает к поверхности сушильного оборудования. В основном из-за трудностей, связанных с обращением с липкой фазой, сушка биошлама в настоящее время считается скорее искусством, чем наукой. Эта статья представляет собой попытку перенести эту деятельность по сушке в область науки путем разработки аналитического подхода к определению начальной и конечной концентрации твердых веществ в зоне липкой фазы биошлама.
Используя в качестве примера данные лабораторных испытаний из предыдущей статьи по этой теме ( Chem. Eng., , сентябрь 2014 г., стр. 51–54), в этой статье представлены схемы проектирования для сушки биошлама, которые могут использовать эти предсказанные начальная и конечная концентрации зоны липкой фазы. Вооружившись информацией, представленной в этой статье, проектировщики и операторы систем сушки биошлама сточных вод могут создавать системы сушки с более высокой эффективностью и меньшим количеством проблем, связанных с липкой фазой.
Границы зоны липкой фазы
Биошлам обычно содержит большое количество воды. Теплотворная способность обычно низкая, и материал трудно использовать непосредственно в качестве топлива. Таким образом, для использования биошлама в качестве топлива необходимо удалить воду перед подачей материала в оборудование для сжигания.
В процессе обезвоживания и сушки осадки сточных вод проходят через так называемую зону липкой фазы. Хотя эта стадия изменения свойств наблюдается давно, впервые количественно она была охарактеризована Петерсом и другими [9].0015 1 ]. В этом направлении исследований использовался лабораторный протокол, основанный на испытаниях на сдвиг, для картирования изменений свойств шлама в процессе сушки. Для получения дополнительной информации о картировании липкости во время сушки биошлама см. Ref. 2.
Хотя эта стадия изменения свойств наблюдается давно, впервые количественно она была охарактеризована Петерсом и другими [9].0015 1 ]. В этом направлении исследований использовался лабораторный протокол, основанный на испытаниях на сдвиг, для картирования изменений свойств шлама в процессе сушки. Для получения дополнительной информации о картировании липкости во время сушки биошлама см. Ref. 2.
В этой статье обсуждались пять различных образцов шлама, которые использовались для тестирования. Эти образцы были взяты на очистных сооружениях Monsanto в Антверпене, Бельгия. Содержание органики в образцах шлама колеблется от 49 до 30 мас. %. Данные испытаний, представленные Петерсом, показывают, что липкие характеристики осадка зависят как от содержания воды в осадке во время сушки, так и от исходного содержания органических веществ. Сводка данных исследования Петерса представлена в таблице 1.
На основании этих данных зона липкой фазы появляется в определенном диапазоне содержания воды. Однако точный диапазон зоны липкой фазы зависит от свойств шлама. Интересно отметить, что чем ниже содержание органики в твердой фракции ила, тем ниже содержание воды в иле, где начинается липкая фаза. Причина того, что высыхающий ил образует липкую фазу, заключается в том, что в процессе биоочистки микроорганизмы захватываются активным илом и образуют длинноцепочечные биополимеры [ 2 ]. Биополимеры «склеивают» микроорганизмы в иле. При обезвоживании и сушке склеивающие биополимеры становятся все более концентрированными, что приводит к образованию липкой смеси.
Однако точный диапазон зоны липкой фазы зависит от свойств шлама. Интересно отметить, что чем ниже содержание органики в твердой фракции ила, тем ниже содержание воды в иле, где начинается липкая фаза. Причина того, что высыхающий ил образует липкую фазу, заключается в том, что в процессе биоочистки микроорганизмы захватываются активным илом и образуют длинноцепочечные биополимеры [ 2 ]. Биополимеры «склеивают» микроорганизмы в иле. При обезвоживании и сушке склеивающие биополимеры становятся все более концентрированными, что приводит к образованию липкой смеси.
Рисунок 1. Свойства липкости осушенного ила с очистных сооружений разделены на три основные зоны
Фазы зоны липкой фазы
В целом, зоны свойств липкости для отдельных образцов ила можно качественно представить на диаграмме показано на рисунке 1, на котором показаны три фазы свойств ила, обычно наблюдаемые во время сушки биошлама. Три этапа описаны далее здесь:
- При высоком содержании воды (низкое содержание твердых веществ) влажный шлам не ведет себя как липкое вещество, поскольку на этой стадии он представляет собой раствор биополимера с низкой концентрацией.

- Когда содержание воды в иле уменьшается (и увеличивается содержание сухих веществ), раствор биополимера становится все более и более концентрированным, а общий биошлам становится все более липким, пока не достигнет зоны липкой фазы. Концентрация и диапазон липкой фазы зависят от свойств ила.
- По мере дальнейшего снижения содержания воды в процессе сушки сразу за пределами зоны липкой фазы липкость осадка резко снижается. Причина в том, что в этот момент биополимеры достаточно сухие, чтобы их можно было диспергировать, не слипаясь.
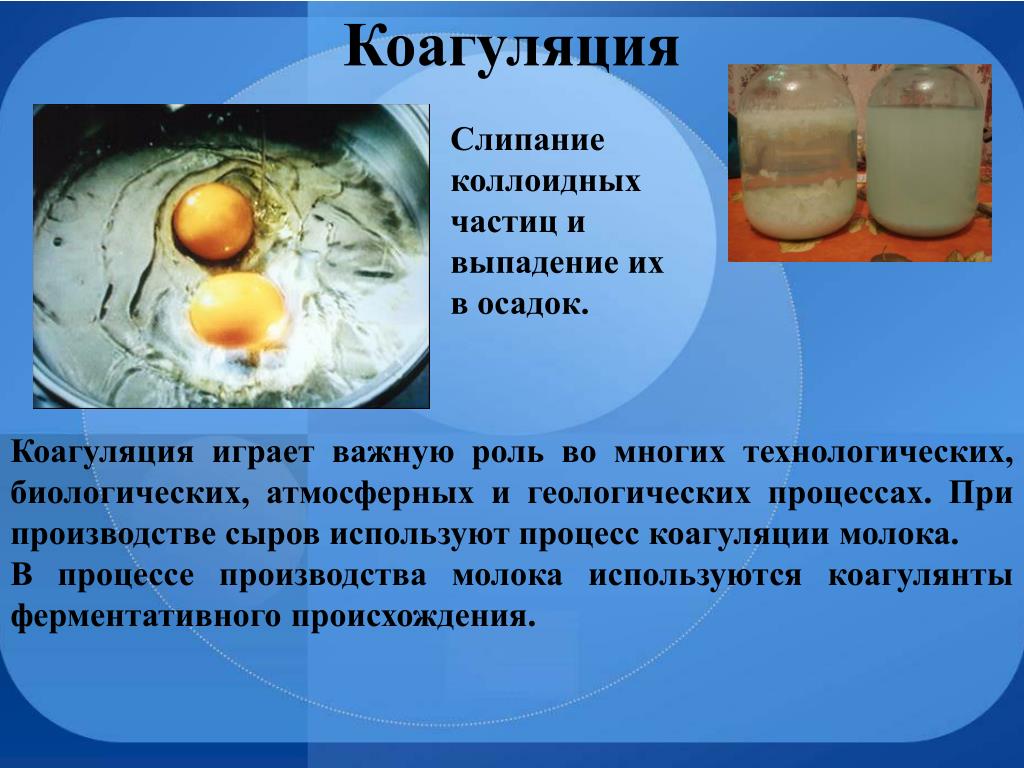
Три этапа процесса сушки осадка показаны на реальных фотографиях на рис. 2. Эти фотографии взяты из фильма о работе сушилки осадка для осадка сточных вод типографии [ 3 ].
Рисунок 2. На серии фотографий показан пример прохождения ила через липкую фазу в процессе сушки
Механизмы вязкой фазы
Почему биошлам с более низким содержанием органических веществ имеет зону вязкой фазы, образующуюся при более низком содержании воды? Более низкое содержание органики в биошламе (в результате временно повышенного количества осажденных солей карбоната кальция в флокуле ила из-за изменения состава сточных вод, например [ 4 ]) предполагает более низкий уровень внеклеточных полимерных веществ на единицу иловая масса. Таким образом, содержание внеклеточных полимерных веществ определяет спектр липкой фазы ила.
Таким образом, содержание внеклеточных полимерных веществ определяет спектр липкой фазы ила.
Прогнозирование начальной и конечной точек
Начальную и конечную концентрации воды в зоне образования липкой фазы ила во время сушки ила можно прогнозировать с помощью двух моделей. Эти модели направлены на то, чтобы исключить некоторые догадки при прогнозировании начальной и конечной точек зоны образования вязкой фазы.
Эти две модели были разработаны с помощью регрессионного анализа на основе данных таблицы 1. Диапазон содержания органических веществ в сухих веществах составляет от 30 до 49 мас.%.
Модель S101 . Данную модель можно использовать для оценки начальной концентрации зоны липкой фазы при сушке биошлама:
% C ч = –62,6563 + 5,638979 % C o – 0,0573 % С или 2 (1)
Модель S102. Данная модель предназначена для оценки конечной концентрации зоны липкой фазы при сушке биошлама:
% C l = 74,707 – 2,5054 % C o + 0,04655 % C o 2 (2)
Где
% C ч весовой процент содержания воды в начале вязкой фазы
% C l весовой процент содержания воды в конце вязкой фазы
% C o – весовое содержание органических материалов в биошламе, поступающем в сушилку осадка.
Коэффициенты корреляции составляют 0,9983 для модели S101 и 0,9699 для модели S102. Ошибки оценки составляют менее ±1% для модели S101 и менее ±5% для модели S102.
Схемы проектирования системы сушки
На этапе проектирования оборудования для процесса сушки осадка существует множество подходов, которые можно использовать для преодоления проблем, связанных с вязкой фазой сушки осадка. Окончательный выбор зависит от требований конкретного применения сушки осадка.
Здесь представлены пять возможных стратегий решения проблем, связанных с липкой фазой, о которых можно узнать, определив начальную и конечную точки липкой фазы, как обсуждалось ранее. Пять стратегий следующие:
Схема A . Высушите осадок до того, как начнется липкая фаза. Если цель сушки осадка состоит только в уменьшении веса и объема, необходимого для транспортировки и утилизации осадка, осадок следует сушить только до момента, когда начинается липкая фаза.
Коэффициент уменьшения высушивания осадка (вес), который представляет собой отношение количества поступающего влажного осадка к расходу сухого осадка, можно рассчитать следующим образом:
R = (100 – % Вт) / % D (3)
Где
R – коэффициент измельчения (масса), скорость поступления влажного ила по сравнению с сухим. производительность
% D весовой процент содержания сухих твердых частиц во влажном шлам
% W весовой процент содержания воды в начале зоны вязкой фазы
Например, для образца ила № 5 в табл. начинается липкая фаза, содержание воды 55%. Коэффициент редукции составляет около 2,25. После того, как шлам в этом образце высушен, необходимая транспортировка и утилизация сокращаются примерно наполовину.
Схема B. Разработайте сушильное оборудование с более высоким крутящим моментом и прочной конструкцией. Этот подход может быть самым дорогим, но он может решить корень проблемы.
Этот подход может быть самым дорогим, но он может решить корень проблемы.
Схема C. Продукты обратного смешивания сухого ила с влажным илом, подаваемым в сушилку, таким образом, чтобы содержание воды в смешанном иле, подаваемом в сушилку, было ниже конечной концентрации липкой фазы. Такой подход увеличит требуемый размер сушилки осадка. Система показана на рис. 3.
Рис. 3. Стратегия сушки, рассмотренная на схеме C, включающая обратное перемешивание сухого ила, показана на этой диаграмме
расход корма (вес) можно рассчитать следующим образом:
R = ( Вт 1 – X ) / ( X – Вт 2 ) (4)
Где
R – отношение между расход рециркулируемого сухого ила и расход подаваемого влажного ила (вес)
W 1 – содержание воды во влажном шламе на входе
W 2 – содержание воды в сухом шламе на выходе
X – содержание воды в шламе в конце зоны липкой фазы.
Схема D. Контролируйте окружающую среду сушилки таким образом, чтобы внутри сушилки содержание воды в высушиваемом осадке всегда было ниже конечной концентрации зоны липкой точки. Такого подхода можно добиться, предварительно заполнив сушильную камеру уже высушенными продуктами. Затем влажный шлам подается в сушилку небольшими порциями за один раз, так что влажный ил легко смешивается с высушенными продуктами, образуя смешанный сыпучий ил. Этот насыпной шлам имеет содержание воды внутри сушилки, которое всегда меньше, чем конечная концентрация зоны липкой фазы.
Схема E. Ввести добавку полиалюминийхлорид (PACl) в подачу влажного шлама для уменьшения или устранения, по возможности, липкости ила [ 5–7 ] Возможность использования PACl для корректировка или кондиционирование влажного ила заключается в том, что связанная гидратационная вода внутри супералюминиевых структур растворов PACl будет прилипать к внешней стороне хлопьев ила. И гидратационная вода действует как смазка, направляя липкие биополимеры через диапазон сухости [ 7 ].
И гидратационная вода действует как смазка, направляя липкие биополимеры через диапазон сухости [ 7 ].
Заключительные замечания
Модели вязкой фазы сушки ила КОС дают оценку начальной и конечной концентраций воды в вязкой фазе. Эти модели представляют собой первоначальную попытку определить диапазон липкой зоны более научно, чем это было возможно в прошлом.
Используя эти предсказанные начальные и конечные концентрации зоны липкой фазы, проектировщики и операторы систем сушки биошлама сточных вод могут избежать некоторых проблем, наблюдаемых с вязкой фазой, и могут разрабатывать системы сушки с более высокой эффективностью. Однако, чтобы сделать модели более полезными, требуется гораздо больше работы по расширению данных для начала и окончания зон вязкой фазы. Данные, использованные в этой статье, основаны на образцах с очистных сооружений Monsanto в Антверпене, Бельгия, но рекомендуется использовать более широкий диапазон образцов, чтобы расширить применимость моделей к биошламам из разных источников и регионов. Эта работа поможет улучшить модели прогнозирования.
Эта работа поможет улучшить модели прогнозирования.
Под редакцией Скотта Дженкинса
Благодарность
Некоторые материалы взяты с разрешения из книги Yen-Hsiung Kiang «Оценка свойств топлива и характеристика процесса горения», опубликованной в 2018 году издательством Academic Press. Это исследование не получило специального грантового финансирования.
Каталожные номера
1. Петерс, Барт, Девил, Раф, Ван Импе, Дж. Ф. М., и Сметс, Илзе, Использование лабораторного протокола на основе испытаний на сдвиг для картирования липкой фазы активного ила, Инженерная экология 28, стр. 81–85, 2011.
2. Питерс, Барт, Девил, Раф, Ван Импе, Дж.Ф.М. и Semts, Ilse, Проблемы сушки липкого осадка сточных вод, Chem. Eng ., сентябрь 2014 г., стр. 51–54.
3. Кианг Йен-Сюн, информация из личной коллекции автора.
4.
 Петерс, Барт, Девил, Раф, Лечата, Д. и Сметс, Илзе, Количественное определение обменного кальция в хлопьях активного ила и его влияние на оседание ила, Технология разделения и очистки , 83, стр. 1–8, 2011.
Петерс, Барт, Девил, Раф, Лечата, Д. и Сметс, Илзе, Количественное определение обменного кальция в хлопьях активного ила и его влияние на оседание ила, Технология разделения и очистки , 83, стр. 1–8, 2011.5. Петерс, Барт, Девил, Раф, Верниммен, Люк и Сметс, Илзе, Предотвращение липкости осадка за счет добавления хлорида полиалюминия (PACl), В: Материалы 11-го Всемирного конгресса по фильтрации (WFC11), Грац, Австрия, статья P368 , 16–20 апреля 2012 г.
6. Петерс, Барт, Девил, Раф и Сметс, Илзе, Усовершенствованное управление технологическим процессом промышленной сушилки осадка с помощью бинарного логистического регрессионного моделирования проблем загрязнения, J. Process Control, 22, стр. 1,387–1,39.6, 2012.
7. Петерс, Барт, Девил, Раф, Верниммен, Люк, Ван ден Богарта, Бенно и Сметс, Илзе, Добавление хлорида полиалюминия (PACl) в отработанный активный ил для смягчения негативного воздействия его липкой фазы в операциях обезвоживания-сушки.
 , Water Research , 47, стр. 3600–3609, 2013.
, Water Research , 47, стр. 3600–3609, 2013.Автор
Йен-Сюн Кианг является техническим директором Xiamen Baipeng Environmental Engineering Co., Ltd. (16F, Yong Tong Chang Building, Jiahe Rd., Xiamen, 36100, Китай; электронная почта: [email: [email] защищено] Телефон: +86-1521-0324628). Кианг имеет более чем 40-летний опыт работы в области топлива, сжигания, сжигания, твердых отходов, сушки осадка сточных вод, компостирования, загрязнения грунтовых вод, загрязнения почвы, когенерации и сжиженного природного газа, очистки сточных вод, загрязнения воздуха и других областях. Он получил степень магистра медицины и докторскую степень. получил степень бакалавра химического машиностроения в Университете Флориды и является зарегистрированным профессиональным инженером в Пенсильвании. Кианг является членом советов директоров Falcon Power Co., Ltd. и Китайской ассоциации альтернативной энергетики, а также консультантом Управления по энергетике и сокращению выбросов углерода Исполнительного юаня Китайской Республики и ведущим консультантом компании Sustainable Development Company, Ltd. На протяжении своей карьеры он выполнял планирование и проектирование сотен центров обработки отходов (включая сжигание, когенерацию, физико-химическую обработку, отверждение и захоронение).
На протяжении своей карьеры он выполнял планирование и проектирование сотен центров обработки отходов (включая сжигание, когенерацию, физико-химическую обработку, отверждение и захоронение).
Руководство пользователя для анализа разделения фаз с использованием очищенных белков
1. Banani S.F., Lee H.O., Hyman A.A., Rosen M.K. Биомолекулярные конденсаты: организаторы клеточной биохимии. Нац. Преподобный Мол. Клеточная биол. 2017;18:285–298. [Бесплатная статья PMC] [PubMed] [Google Scholar]
2. Хайман А.А., Вебер К.А., Юлихер Ф. Разделение фаз жидкость-жидкость в биологии. Анну. Преподобный Cell Dev. биол. 2014; 30:39–58. [PubMed] [Google Scholar]
3. Shin Y., Brangwynne C.P. Конденсация жидкой фазы в физиологии клетки и болезни. Наука. 2017;357 [PubMed] [Google Scholar]
4. Нотт Т.Дж., Петсалаки Э., Фарбер П., Джервис Д., Фусснер Э., Плоховиц А., Крэггс Т.Д., Базетт-Джонс Д.П., Поусон Т., Форман-Кей Дж.Д., Болдуин А.Дж. Фазовый переход неупорядоченного белка nuage приводит к образованию экологически чувствительных безмембранных органелл. Мол. Клетка. 2015;57:936–947. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Мол. Клетка. 2015;57:936–947. [Бесплатная статья PMC] [PubMed] [Google Scholar]
5. Li P., Banjade S., Cheng H.-C., Kim S., Chen B., Guo L., Llaguno M., Hollingsworth J.V., Кинг Д.С., Банани С.Ф., Руссо П.С., Цзян К.-С., Никсон Б.Т., Розен М.К. Фазовые переходы в сборке поливалентных сигнальных белков. Природа. 2012; 483:336–340. [Бесплатная статья PMC] [PubMed] [Google Scholar]
6. Патель А., Ли Х.О., Яверт Л., Махарана С., Джанел М., Хайн М.Ю., Стойнов С., Махамид Дж., Саха С., Францманн Т.М., Позняковски А., Позер И., Магелли Н., Ройер Л.А., Вейгерт М., Майерс Э.В., Грилл С., Дрексел Д., Хайман А.А., Альберти С. Фазовый переход белка FUS БАС из жидкого состояния в твердое, ускоренный мутацией заболевания. Клетка. 2015; 162:1066–1077. [PubMed] [Google Scholar]
7. Саха С., Вебер К.А., Ноуш М., Адаме-Арана О., Хёге К., Хайн М.Ю., Осборн-Нишимура Э., Махамид Дж., Джахнел М., Джаверт Л., Позняковски А., Экманн К.Р., Юлихер Ф., Хайман А.А. Полярное расположение разделенных фаз жидких компартментов в клетках регулируется механизмом конкуренции мРНК. Клетка. 2016 [Бесплатная статья PMC] [PubMed] [Google Scholar]
Клетка. 2016 [Бесплатная статья PMC] [PubMed] [Google Scholar]
8. Wang J.T., Smith J., Chen B.-C., Schmidt H., Rasoloson D., Paix A., Lambrus B.G., Calidas D., Betzig E., Seydoux G. Регуляция динамики гранул РНК с помощью фосфорилирование богатых серином внутренне неупорядоченных белков в C. elegans . жизнь. 2014;3 [бесплатная статья PMC] [PubMed] [Google Scholar]
9. Holehouse A.S., Pappu R.V. Белковые полимеры: кодирование фазовых переходов. Нац. Матер. 2015;14:1083–1084. [PubMed] [Google Scholar]
10. Альберти С., Хайман А.А. Являются ли аберрантные фазовые переходы движущей силой клеточного старения? Биоэссе. 2016;38:959–968. [Бесплатная статья PMC] [PubMed] [Google Scholar]
11. Rabouille C., Alberti S. Адаптация клеток при стрессе: новая роль безмембранных компартментов. Курс. мнение Клеточная биол. 2017;47:34–42. [PubMed] [Google Scholar]
12. Seydoux G., Braun R.E. Путь к тотипотентности: уроки зародышевых клеток. Клетка. 2006; 127:891–904. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
13. Кнутсон А.К., Эгельхофер Т., Рехтштайнер А., Строум С. Зародышевые гранулы предотвращают накопление соматических транскриптов у взрослых Caenorhabditis elegans Зародышевая линия. Генетика. 2017; 206: 163–178. [Бесплатная статья PMC] [PubMed] [Google Scholar]
14. Апдайк Д.Л., Кнутсон А.К., Эгельхофер Т.А., Кэмпбелл А.С., Строум С. Компоненты зародышевых гранул предотвращают соматическое развитие в зародышевой линии C. elegans . Курс. биол. 2014;24:970–975. [PMC free article] [PubMed] [Google Scholar]
15. Воронина Е., Сейду Г., Сассоне-Корси П., Нагамори И. Гранулы РНК в половых клетках. Харб Колд Спринг. Перспектива. биол. 2011;3 [Бесплатная статья PMC] [PubMed] [Google Scholar]
16. Updike D., Strome S. Сборка и функция гранул P в зародышевых клетках Caenorhabditis elegans . Дж. Андрол. 2010; 31:53–60. [Бесплатная статья PMC] [PubMed] [Google Scholar]
17. Kawasaki I., Shim Y.H., Kirchner J. , Kaminker J., Wood W.B., Strome S. PGL-1, предсказанный РНК-связывающий компонент зародышевых гранул , необходим для фертильности C. elegans . Клетка. 1998; 94: 635–645. [PubMed] [Google Scholar]
, Kaminker J., Wood W.B., Strome S. PGL-1, предсказанный РНК-связывающий компонент зародышевых гранул , необходим для фертильности C. elegans . Клетка. 1998; 94: 635–645. [PubMed] [Google Scholar]
18. Kawasaki I., Amiri A., Fan Y., Meyer N., Dunkelbarger S., Motohashi T., Karashima T., Bossinger O., Strome S. Белки семейства PGL связываются с зародышевыми гранулами и функционируют избыточно в Caenorhabditis elegans Развитие зародышевой линии. Генетика. 2004; 167: 645–661. [PMC free article] [PubMed] [Google Scholar]
Множественные потенциальные геликазы зародышевой линии являются компонентами специфичных для зародышевой линии P-гранул Caenorhabditis elegans . проц. Натл. акад. науч. США, 1996; 93:13837–13842. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Комбинаторная РНК-интерференция указывает на то, что GLH-4 может компенсировать GLH-1; эти два компонента гранул P имеют решающее значение для фертильности в С. Элеганс . Разработка. 2000;127:2907–2916. [PubMed] [Google Scholar]
Разработка. 2000;127:2907–2916. [PubMed] [Google Scholar]
21. Spike C.A., Bader J., Reinke V., Strome S. DEPS-1 способствует сборке P-гранул и интерференции РНК в зародышевых клетках C. elegans . Разработка. 2008; 135:983–993. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Гранулы Germline P представляют собой жидкие капли, которые локализуются путем контролируемого растворения/конденсации. Наука. 2009 г.;324:1729–1732. [PubMed] [Google Scholar]
23. Conduit P.T., Wainman A., Raff J.W. Функция и сборка центросом в клетках животных. Нац. Преподобный Мол. Клеточная биол. 2015;16:611–624. [PubMed] [Google Scholar]
24. Вудрафф Дж.Б., Вуэсеке О., Хайман А.А. Структура и динамика перицентриолярного материала. Филос. Транс. Р. Соц. Лонд. сер. Б биол. науч. 2014;369 [бесплатная статья PMC] [PubMed] [Google Scholar]
25. Вудрафф Дж.Б., Феррейра Гомес Б., Видлунд П.О., Махамид Дж., Хонигманн А., Хайман А.А. Клетка. 1 июня 2017 г .;169(6): 1066–1077. e10. [PubMed] [Google Scholar]
e10. [PubMed] [Google Scholar]
26. Вудрафф Дж.Б., Вуэсеке О., Вискарди В., Махамид Дж., Очоа С.Д., Бункенборг Дж., Видлунд П.О., Позняковский А., Занин Э., Бахманьяр С., Зинке А. ., Хонг С.Х., Декер М., Баумайстер В., Андерсен Дж.С., Огема К., Хайман А.А. центросомы. Регулируемая сборка надмолекулярного центросомного каркаса in vitro. Наука. 2015; 348: 808–812. [Бесплатная статья PMC] [PubMed] [Google Scholar]
27. Гелл С., Фрил С.Т., Боргоново Б., Дрексель Д.Н., Хайман А.А., Ховард Дж. Очистка тубулина из мозга свиньи. Методы Мол. биол. 2011; 777:15–28. [PubMed] [Академия Google]
28. Альберти С., Хальфманн Р., Кинг О., Капила А., Линдквист С. Систематический обзор идентифицирует прионы и освещает особенности последовательности прионогенных белков. Клетка. 2009; 137: 146–158. [PMC free article] [PubMed] [Google Scholar]
29. Mukhopadhyay S., Krishnan R., Lemke E.A., Lindquist S., Deniz A.A. Естественно развернутый мономер дрожжевого приона принимает ансамбль коллапсированных и быстро флуктуирующих структур. проц. Натл. акад. науч. США 2007;104:2649–2654. [Бесплатная статья PMC] [PubMed] [Google Scholar]
проц. Натл. акад. науч. США 2007;104:2649–2654. [Бесплатная статья PMC] [PubMed] [Google Scholar]
30. Серио Т.Р., Кашикар А.Г., Коваль А.С., Савицкий Г.Дж., Мослехи Дж.Дж., Серпелл Л., Арнсдорф М.Ф., Линдквист С.Л. Конформационное преобразование ядра и репликация конформационной информации прионной детерминантой. Наука. 2000; 289:1317–1321. [PubMed] [Google Scholar]
31. Гловер Дж.Р., Коваль А.С., Ширмер Э.К., Патино М.М., Лю Дж.Дж., Линдквист С. Самозасеваемые волокна, образованные Sup35, белковой детерминантой [PSI +], наследуемой прион- подобный фактор S. cerevisiae . Клетка. 1997; 89: 811–819. [PubMed] [Google Scholar]
32. Кинг О.Д., Гитлер А.Д., Шортер Дж. Верхушка айсберга: РНК-связывающие белки с прионоподобными доменами при нейродегенеративных заболеваниях. Мозг Res. 2012;1462:61–80. [Бесплатная статья PMC] [PubMed] [Google Scholar]
33. Малиновская Л., Крошвальд С., Альберти С. Белковые расстройства, склонность к прионам и самоорганизующиеся макромолекулярные коллективы. Биохим. Биофиз. Акта. 2013; 1834: 918–931. [PubMed] [Академия Google]
Биохим. Биофиз. Акта. 2013; 1834: 918–931. [PubMed] [Академия Google]
34. Krammer C., Kryndushkin D., Suhre M.H., Kremmer E., Hofmann A., Pfeifer A., Scheibel T., Wickner R.B., Schätzl H.M., Vorberg I. Дрожжевой домен Sup35NM размножается как прион в клетках млекопитающих. . проц. Натл. акад. науч. США 2009;106:462–467. [Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Серио Т.Р., Кашикар А.Г., Мослехи Дж.Дж., Коваль А.С., Линдквист С.Л. Дрожжевой прион [psi +] и его определитель Sup35p. Методы Энзимол. 1999; 309: 649–673. [PubMed] [Академия Google]
36. Шейбель Т., Линдквист С.Л. Роль конформационной гибкости в размножении и поддержании прионов для Sup35p. Нац. Структура биол. 2001; 8: 958–962. [PubMed] [Google Scholar]
37. Franzmann T.M., Jahnel M., Pozniakovsky A., Mahamid J., Holehouse A.S., Nüske E., Richter D., Baumeister W., Grill S.W., Pappu R.V., Hyman A.A., Альберти С. Фазовое разделение дрожжевого прионного белка способствует приспособленности клеток. Наука. 2018;359 [PubMed] [Google Scholar]
Наука. 2018;359 [PubMed] [Google Scholar]
38. Molliex A., Temirov J., Lee J., Coughlin M., Kanagaraj A.P., Kim H.J., Mittag T., Taylor J.P. Разделение фаз доменами низкой сложности способствует стрессу сборка гранул и приводит к патологической фибрилляции. Клетка. 2015; 163:123–133. [Бесплатная статья PMC] [PubMed] [Google Scholar]
39. Лин Ю., Проттер Д.С.В., Розен М.К., Паркер Р. Формирование и созревание капель жидкости, разделенных фазами, с помощью РНК-связывающих белков. Мол. Клетка. 2015;60:208–219. [Бесплатная статья PMC] [PubMed] [Google Scholar]
40. Мураками Т., Камар С., Лин Дж. К., Ширле Г. С. К., Рис Э., Мияшита А., Коста А. Р., Додд Р. Б., Чан Ф. Т. С., Мишель Ч. Х., Кроненберг-Верстег Д., Ли Ю., Ян С.-П., Вакутани Ю., Медоуз В., Ферри Р.Р., Донг Л., Тарталья Г.Г., Фаврин Г., Лин В.-Л., Диксон Д.В., Жень M., Ron D., Schmitt-Ulms G., Fraser P.E., Shneider N.A., Holt C., Vendruscolo M., Kaminski C.F., St George-Hyslop P. Индуцированный мутацией ALS/FTD фазовый переход жидких капель FUS и обратимый гидрогелей в необратимые гидрогели ухудшает функцию гранул РНП. Нейрон. 2015; 88: 678–690. [PMC free article] [PubMed] [Google Scholar]
Нейрон. 2015; 88: 678–690. [PMC free article] [PubMed] [Google Scholar]
Альберти С. Аберрантный фазовый переход стрессовых гранул, вызванный неправильно свернутым белком и предотвращенный функцией шаперона. EMBO J. 2017; 36: 1669–1687. [Бесплатная статья PMC] [PubMed] [Google Scholar]
42. Маккензи И.Р., Николсон А.М., Саркар М., Мессинг Дж., Пьюрис М.Д., Потье К., Анну К., Бейкер М., Перкерсон Р.Б., Курти А. ., Матчетт Б.Дж., Миттаг Т., Темиров Дж., Хсюн Г.-Ю.Р., Кригер К., Мюррей М.Е., Като М., Фрайер Дж.Д., Петручелли Л., Зинман Л., Вайнтрауб С., Месулам М., Кит Дж., Живкович С.А., Хирш-Райншаген В., Роос Р.П., Зюхнер С., Графф-Рэдфорд Н.Р., Петерсен Р.К., Казелли Р.Дж., Всолек З.К., Фингер Э., Липпа С., Лакомис Д., Стюарт Х., Диксон Д.В., Ким Х.Дж., Рогаева Э., Бигио Э., Бойлан К.Б., Тейлор Дж.П., Радемакерс Р. Мутации TIA1 при боковом амиотрофическом склерозе и лобно-височной деменции способствуют разделению фаз и изменению динамики стрессовых гранул. Нейрон. 2017;95:808–816. e9. [Бесплатная статья PMC] [PubMed] [Google Scholar]
e9. [Бесплатная статья PMC] [PubMed] [Google Scholar]
43. Ван Дж., Чой Дж.М., Холехаус А.С., Ли Х.О., Чжан С., Джахнел М., Махарана С., Леметр Р., Позняковский А., Дрексель Д., Посер И., Паппу Р.В., Альберти С., Хайман А.А. Молекулярная грамматика, лежащая в основе движущих сил разделения фаз прионоподобных РНК-связывающих белков. Клетка. 2018 [Бесплатная статья PMC] [PubMed] [Google Scholar]
44. Боке Э., Руер М., Вюр М., Кафлин М., Леметр Р., Гиги С.П., Альберти С., Дрексель Д., Хайман А.А. , Митчисон Т.Дж. Амилоидоподобная самосборка клеточного компартмента. Клетка. 2016; 166: 637–650. [Бесплатная статья PMC] [PubMed] [Google Scholar]
45. Ланкастер А.К., Наттер-Апхам А., Линдквист С., Кинг О.Д. PLAAC: веб-приложение и приложение командной строки для идентификации белков с прионоподобным аминокислотным составом. Биоинформатика. 2014;30:2501–2502. [Бесплатная статья PMC] [PubMed] [Google Scholar]
46. Dosztányi Z., Csizmok V., Tompa P., Simon I. IUPred: веб-сервер для прогнозирования внутренне неструктурированных областей белков на основе расчетного содержания энергии.